Notre organisme, et particulièrement nos muscles, a besoin d'énergie pour fonctionner. Cette énergie est fournie par les nutriments. Cependant une utilisation directe est impossible dans l'organisme. Tout transfert d'énergie passe par l'intermédiaire d'une molécule appelée ATP (adénosine tri-phosphate) :
Une adénosine tri-phosphate contient 3 molécules de phosphate riche en énergie. Lorsqu'on détache le 3ème phosphate, on libère ainsi une grande quantité d'énergie. La molécule ainsi obtenue est l'ADP (adénosine di-phosphate). La libération du 3ème phosphate se fait par hydrolyse et l'accrochage d'un phosphate à une molécule est appelée phosphorylation. Rappelons qu'un phosphate isolé sera ainsi un phosphate inorganique Pi. La réaction est la suivante :
 Comment la cellule musculaire fabrique de l'ATP
Comment la cellule musculaire fabrique de l'ATP
L'activité physique est conditionnée par l'activité des cellules musculaires. Comme toute cellule, la cellule musculaire (ou fibre musculaire) a besoin d'énergie, c'est à dire d'ATP, pour fonctionner.

L'activité physique est conditionnée par l'activité des cellules musculaires. Comme toute cellule, la cellule musculaire (ou fibre musculaire) a besoin d'énergie, c'est à dire d'ATP, pour fonctionner.
Lorsqu'on parle d'une activité musculaire, il faudrait plutôt parler des activés musculaires. En effet, selon que l'on soulève un poids ou que l'on fait un jogging, l'effort n'est pas le même.
Le tableau suivant, met en évidence, ces différences à partir de quelques sports :

L'effort d'un haltérophile est bref et très demandeur en énergie. C'est un sport qui demande de l'explosivité. L'ATP nécessaire à cet effort est fabriqué à partir d'une molécule phosphorylée, la phosphocréatine (P-Cr) comme le montre le graphe suivant :


On constate que la réaction ne nécessite pas de dioxygène. On est donc dans une réaction anaérobie. Cette réaction ne produit pas de substance de déchet comme l'acide lactique. On parle de métabolisme anaérobie alactique.
La créatine est une molécule synthétisée par le corps humain. Elle est cependant utilisée comme supplément par le sportifs (et les sportifs du dimanche !) avec des résultats extrêmement aléatoire, selon l'individu et la qualité du produit.

Si un effort explosif demande un métabolisme particulier, il sollicite aussi des fibres musculaires particulières. Les tableaux suivants précisent les spécificités métaboliques des fibres musculaires :


On constate que les fibres de type II possèdent un métabolisme anaérobie alactique. Ce sont donc ces fibres II qui sont le plus sollicitées en haltérophilie et dans les sports explosifs.

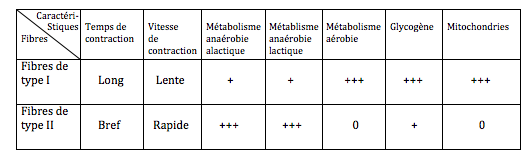
On constate que les fibres de type II possèdent un métabolisme anaérobie alactique. Ce sont donc ces fibres II qui sont le plus sollicitées en haltérophilie et dans les sports explosifs.
Si on revient au tableau des différents sports, on constate que lors d'un effort de quelques minutes, le métabolisme de la cellule musculaire n'est pas le même. Il y a production d'acide lactique comme le montre le document suivant :

On constate que dès que le taux de phosphocréatine a atteint un niveau assez bas, la production d'acide lactique se met a grimper.
On peut indiquer la réaction qui forme l'acide lactique musculaire :


Il existe dans les tissus des formes de stockage du glucose. Une est particulièrement important, le glycogène.

Nous allons donc vérifier qu'il existe des réserve de glycogène dans le muscle, en utilisant de la viande de bœuf.
Protocole

On obtient le résultat suivant :

On obtient aussi une précipitation du glycogène dans l'éthanol.
Le glycogène musculaire, par glycogénolyse, va donc donner des molécules de glucose capables de fournir le substrat nécessaire à la réaction. Ce sont aussi les fibres de type II qui assurent le métabolisme anaérobie lactique.
Dans le cas des sports de fond, c'est le métabolisme aérobie qui prend la relève. On est dans le cadre de la respiration cellulaire. C'est toujours le glycogène qui est la source de l'énergie mais dans ce cas, après glycogénolyse, le glucose est totalement oxydé en CO2.
Utilisation du glycogène chez un marathonien.

Dans ce cas, comme l'indique le tableau déjà cité, ce sont les fibres de type I qui sont sollicitées.


Nous venons de voir comment la cellule musculaire récupère de l'énergie sous forme d'ATP.
Intéressons nous maintenant aux mécanismes de dépense d'énergie, c'est à dire à la contraction musculaire.
Un muscle est constitué de grandes cellules contenant plusieurs noyaux, appelées fibres musculaires ou myocytes. Le cytoplasme est appelée sarcoplasme. La cellule musculaire contient des fibrilles protéiques appelées myofibrilles.

Les documents suivants montrent la structures des fibres musculaires :



les myofibrilles sont essentiellement constitués d'actine et de myosine. Leurs structures sont données sur le document ci-dessous :

La structure des fibrilles varie selon que la fibre est contractée ou décontractée.

La contraction musculaire est due à la rotation des têtes de myosine qui s'accrochent au filaments d'actines. Cet accrochage permet de déplacement relatif des fibres ce qui entraîne la contraction musculaire.
Le mouvement de la tête de myosine nécessite de l'énergie. c'est à ce niveau que l'ATP fabriqué par la cellule musculaire va être consommé selon les modalités suivantes :

Le mouvement de la tête de myosine nécessite de l'énergie. c'est à ce niveau que l'ATP fabriqué par la cellule musculaire va être consommé selon les modalités suivantes :


















